 |
 |
 |
 |
Running Title: Individual-based, spatially explicit behavior and food web complexity
Key Words: food web complexity, trophic interactions, spatially-explicit dynamics, individual-based behavior, ecological simulator.
Corresponding Author: Oswald J. Schmitz, Yale University, School of Forestry and Environmental Studies, 370 Prospect St., New Haven, CT 06511 (203) 432-5110; e-mail schmitz@minerva.cis.yale.edu.
Summary
We present a prototype simulator that enables one to explore the influence of individual behavior on the dynamics and structural complexity of food webs. In the simulations, individuals act according to simple, biologically plausible rules in a spatially explicit setting. We present the results of a series of simulation experiments on artificial, 3-trophic level food chains used to calibrate the simulator against real-world systems and to demonstrate the simulator's promise for ecological modeling. Our primary objective was to discover the biological features leading to stability of artificial food chains over ecological time and under different conditions of trophic efficiency. This involved a qualitative analysis of food chains comprised of a plant, herbivore and carnivore species. We explored the consequences of allowing individual heterotrophs to make active choices about resource selection (perception and intentional behavior) under high and low degrees of trophic efficiency. We found that individuals had to adopt realistic behavioral ecological strategies, such as active resource selection, in order for systems to persist, especially under conditions in which trophic efficiencies were of the magnitude observed in real systems (e.g. 10%). Our results reaffirm previous convictions that a better understanding of food web interactions in real-world systems will require approaches that blend animal behavioral ecology with population and community ecology. However, the evidence comes from a new mathematical perspective.
Introduction
It is self-evident in ecology that there is a high degree of complexity both in the organizational structure and dynamics of food webs. Food webs contain large numbers of species interacting via complex networks of direct and indirect pathways (Schoener, 1993; Yodzis, in press). The interactions also take place among different organizational levels and spatial scales (Levin, 1992; McCauley et al., 1993; Brown, 1994). For example, individual physiology and behavioral ecology may have a profound influence on interactions occurring at the level of populations and communities (Holt, 1983; Abrams, 1990; 1993; Fryxell and Lundberg, 1994; Schmitz, 1995). The key, then, to making sense of the daunting complexity is to develop theory on food web dynamics that formalizes how processes at one organizational scale are linked to other organizational scales.
In this paper, we present a prototype simulator which attempts to model food web complexity directly by defining behavioral rules for individual members of a species and using those rules to drive simulations which explore dynamics of the species community as a whole. We show that individuals acting according to rules based on a realistic set of biological primitives on a spatially explicit, yet flexible landscape can have an important influence on the longterm stability of a food web. Our results reaffirm previous convictions (Holt, 1983; Ives and Dobson, 1987; Abrams, 1990; 1993; Wootton and Power, 1993; Fryxell and Lundberg, 1994; Seymour, 1994; Schmitz, 1995) that a better understanding of food web interactions in real-world systems will require approaches that blend animal behavioral ecology with population and community ecology.
One of the most significant hurdles in developing multiscale theories of food web dynamics is limitations imposed by Lotka-Volterra and related differential equation models. Such models are based on assumptions that individuals interact according to mass action kinetics and, in most cases, that affects on individuals in a population are distributed in an egalitarian manner. These assumptions produce models that are largely phenomenological with regard to within and between species interactions thereby limiting the degree of mechanistic understanding of natural systems. Moreover, differential equation models become difficult to solve analytically for systems involving more than 5 species, so, our understanding of dynamics in complex species assemblages usually must be obtained numerically (e.g. see Post and Pimm, 1983; Yodzis 1988, Drake 1990). It is no coincidence that the bulk of community ecological theory that attempts to capture the biological processes in any reasonable detail typically explores dynamical interactions among, at most, three species.
Recent advances in computer technology has been a strong impetus for the development of computational approaches in ecological theory development. These new approaches explore dynamics in an individual-based, spatially explicit setting. In most models, individuals interact on a geography (usually a square lattice either with absorbing or periodic boundary conditions), under a set of rules of engagement based on individual state which determine resource acquisition and reproduction. Typically, one finds that interactions among organisms at one level of organization can have an important influence on the dynamical characteristics of higher organizational levels (Hastings, 1990; 1993; McLaughlin and Roughgarden, 1991; Kim, 1992; DeAngelis et al., 1993; McCauley et al., 1993; Wilson et al., 1993; Durrett and Levin, 1994). For instance, diffusion of individuals in space can cause an otherwise chaotic system to undergo periodic dynamics (e.g. Hastings 1993). Local mobility of predators and prey cause unique patterns of patch structure to emerge over space (Wilson et al. 1993).
Most lattice-based simulators to date explore dynamics in one or two species systems (e.g. McLaughlin and Roughgarden, 1991; Kim, 1992; McCauley et al., 1993; Wilson et al., 1993; Durrett and Levin, 1994). However, it may be difficult to move to greater species complexity using simulations on square (two-dimensional) lattices. This is because of an unrealistic feature of square lattices that individuals could be surrounded by a surfeit of resources yet be unable to reproduce because there is no empty grid position that offspring could occupy. In essence, lattices impose limitations on neighborhood definitions that limit the number of individuals and therefore different species that could exist in any one location. This artificially limits the degree of interactivity among individuals of different species. Consequently, it will likely be very difficult to explore complexity beyond 3-species systems in any reasonable, realistic detail using two dimensional lattice-based approaches.
We approach individual-based spatially explicit modeling by expanding the Echo simulator conceptualized by Holland (1992, 1994). Echo employs a closed-packed ordering of individuals in space which allows offspring to be inserted next to their parents regardless of neighborhood occupancy. Echo is a generic simulator designed to explore interactions among large numbers of different agents. Agents here are a proxy for individuals of a species. Agents have explicit phenotypes which embody artificial chromosomes that code the rules of interaction. Collections of agents interact in space according to their phenotype. Echo offers the possibility to explore the consequences of a rich variety of animal behaviors on trophic interactions. The potential for Echo to offer a better understanding of dynamics in complex ecological systems has been recognized (Brown, 1994; Gell-Mann, 1994; Holland, 1994), but, the extent to which it explains real-world dynamics is largely undetermined (Mitchell and Forrest, 1994).
The primary purpose of this paper is to introduce a prototype version of the Echo-type simulator that we developed to explore the effects of behavioral ecology on species interactions in ecological food webs. We conducted a series of simulation experiments on artificial, 3-trophic level food chains comprised of a plant, herbivore and carnivore species. We found that persistence of complex organizational structure (i.e. full 3-level chains) when trophic efficiencies were of the magnitude observed in real systems (i.e. 10-30%) was only possible if heterotrophic agents could perceive and intentionally select the most nutritious prey agents within their neighborhood.
Description of the simulator
Echo was designed to explore how simple rules could lead to complex organizational structure in a variety of systems. The primary focus was to explore systems such as global economies in which trade (bilateral resource exchange) was a vital interaction. The version of Echo described by Holland (1992) deliberately abstracts away many of the mechanistic details of real systems to concentrate on basic agent-agent and agent-resource interactions. This has given Echo greater generality than traditional numerical computer simulators that tend to be data voracious. But, this comes at the expense of quantitative predictive power for specific systems. In essence, Echo was designed to facilitate "gedanken" experiments (Holland 1992) in order to understand process. As such, it was not immediately applicable to exploring food web dynamics and complexity in any realistic ecological detail.
We have, therefore, modified Echo extensively to go beyond mere thought experiments and allow for more realistic simulation of biological and ecological processes. Our version of Echo, called Gecko, is an ecological simulator that is a simplification of the version described by Holland (1992) with regards to the range of interactions among agents. But, it is an elaboration with regard to resource transformations, timing and identifying how agents interact and with whom. The inspiration for these modifications was to (1) model better resource flows through food webs, (2) allow for unilateral resource flows (herbivory, parasitism and carnivory) instead of primarily bilateral resource flow (i.e., trade), (3) incorporate adaptive (intentional) behavior of individuals, and (4) allow for better adjustment of time scales for interaction among different kinds of agents in different trophic levels.
Gecko, like its progenitor Echo, is essentially a game in which individual agents try to make the most of their environment, defined by site and world definitions and the complex adaptive environment created by the action of other agents in that same environment. The simulator is hierarchical in that: (1) the capacity for interaction is coded at the individual agent level, (2) resource production is specified at each site, and (3) available resources and general rules specifying the magnitude of resource exchange among agents for all sites is specified at a global level.
The basic Gecko topology is presented in Fig. 1. The simulation environment is potentially composed of several sites with differing resource production, but all work presented here concentrates on dynamics in a single-site world. It should be noted that site, here, is not equivalent to a grid coordinate on a discrete lattice. Rather, a site is analogous to a field study site in which species interact locally. Agents within a site have a cyclic, one-dimensional ordering. \Each agent has a specified movement range, not necessarily the two adjacent positions. New agents are inserted into this order next to their parents, dead agents are removed, and the end of the list circles back to the beginning so there are no empty spaces (i.e. a closed-packed ordering). This removes the unrealistic feature of lattice models that an agent could be surrounded by a surfeit of resources, yet be unable to reproduce because there is no empty grid position that offspring could occupy. In effect "crowding" in our simulator is endogenously defined by the balance between resource supply and exploitation by consumers. So, density-dependence is manifest as consumptive competition rather than as interference competition for locations in space. We have to ability to include interference competition. But, this interaction would be coded in the combat matrix (see below) which assigns resource pay-offs to individuals based on the agent with whom they interact (e.g. prey agent, competitor agent etc.).
The disadvantage of the cyclic ordering is that it sacrifices some of the two dimensional qualities of lattices. For example, with only one spatial dimension available to agents, standing waves can form and travel around the circle. This simulation artifact is not essentially different in kind from those caused by arbitrary "crowding" on a discrete grid. Once we identify agent and environmental characteristics leading to this artifact we can treat those characteristics as limits for our simulations in parameter space. Insofar as we are only interested in qualitative system dynamics that depend on the character of local interactions, the linear ordering is sufficient to simulate the effects of spatial dynamics.
The site determines the magnitude of resource influx in each round of simulation. Resource types are coded using a finite alphabet (i.e., a, b, c, d, etc.). Agents present on a site consume resources either from the site (i.e., autotrophs) or from other agents at the site (i.e., heterotrophs).
Agent description
Agents are individuals which may interact with other individuals, are capable of reproduction and subject to mortality from a variety of causes. The strategies or capacities of an agent are specified by a small set of strings or "chromosomes" defined using the same characters as in the resource alphabet (i.e. a, b, c, d, etc.). There is a one to one mapping of the genetic make-up of an agent and the resources it exploits.
In Echo, there are two classes of chromosomes in each agent that determine the exploitation strategy for resources. The first, tag chromosomes, determine the agent's external phenotypic characteristics. There are three tag chromosomes (1) offense tag, (2) defense tag, and (3) mating tag (defining species and sex recognition). Tags are displayed on the exterior of an agent and are analogous to signature groups of an antigen or the logo of an organization (Holland 1992). The second class of chromosomes are condition chromosomes: (1) combat condition, (2) trading condition and (3) mating condition. Conditions determine what kinds of interactions take place when agents encounter one another.
In Gecko, we have consolidated the basic chromosome definitions. An agent has four chromosomes that define interactions: tag, condition, offense and defense. We have also added 2 additional chromosomes, transform from and transform to, to specify resource transformations. The tag string is an agent's external marking. Other agents register this tag and, based on it, decide whether or not to interact with this agent. They do this with their condition string. If agent A's condition matches agent B's tag in whole or in part then agent A will "want" to interact with B. The actual interaction is controlled by the offense and defense strings. Once an interaction is instigated by one agent, each agent's offense string is pitted against the other's defense string to specify the kind and amount of resources that are transferred from one agent to another. This could range from one agent killing the other unilaterally, (or both agents being killed), to a mutually beneficial trade, or somewhere along this continuum (e.g., herbivory resulting in plants unilaterally losing some of their biomass but not being killed). The exact resource exchange will depend on the offense and defense strings and the scoring function for resource exchange (see description of the combat matrix below).
In addition to the chromosome strings, Gecko agent descriptions contain explicit constraints on resource uptake, resource transformation, maintenance metabolic costs, sex and mating. The basic clock cycle in Gecko is the "round" which may be equivalent to a complete generation or fraction thereof depending on the agent. Agents can instigate any number of consumer-resource interactions with other agents per round depending on the level of interactivity specified. For instance, setting interactivity greater than one allows an agent to seek prey more than once per round. However, an agent may only consume a fixed amount of resources within a round. This limit is imposed on raw resource uptake (consumer satiation) not on resource transformation. Gecko imposes a maintenance tax once per round. This is analogous to a maintenance metabolic cost. If an agent does not have sufficient resources to meet metabolic costs it dies.
Agents are eligible to reproduce once they acquire a prespecified multiple of the number of resources and resource types needed to copy their entire genetic make-up (i.e., tag, condition, offense, defense and transform strings). This allows for resource reserves so that agents survive reproduction. The important feature is that an agent's ability to survive and reproduce is explicitly determined by its physiological condition and depends on resource acquisition rates.
In Gecko, we have added resource transformation which allows agents to create new resources from those that are consumed. This makes resource utilization a value-added interaction rather than simply an exchange of resources. Resource transformation is analogous to a heterotroph converting energy from plants into animal protein or a secondary consumer converting one type of animal protein into another type. Resource transformation is also not a one to one conversion hence we build in a trophic efficiency. A plant agent may accumulate resources of one type, say a (sunlight) which it converts to b (plant carbon). Upon eating a plant, a herbivore must convert resource b into resource c (herbivore protein) to copy its own genetic make-up. We build in a trophic efficiency by specifying that a consumer requires n units of resource a, b etc. before it can create one unit of consumer biomass (b, c etc.).
Agents existing in a site are ordered along a one-dimensional ring. Agents instigating an interaction move next to target agents according to movement rules that are coded in the simulator (see below). Reproduction can occur parthenogenically or through mating. Mating is most likely to occur among nearest neighbors of the same species (same tag), if it is enabled. In all cases of reproduction, newborns are inserted next to their parents. Movement to instigate consumer-resource interactions can be varied depending on the degree of behavioral complexity one wishes to incorporate (e.g. resource selection).
Site description
Each site is assigned a function which specifies the influx of resources to the site per round. For example, in the case where resource a (sunlight or mineral nutrients) is supplied at a fixed rate to the site, but resources b, c, d, etc. are only obtained through consumer-resource interactions and resource transformation, the function for a will have a positive value whereas the function for the other resources will have value 0. In Gecko, it is also possible to specify a maximum amount of resources supplied to the site that may persist freely in the environment each round. This level is independent of the maximum allowable uptake of each agent. Consequently, it is possible to vary the degree of resource limitation in the simulations. Also, in each round, a certain fraction of agents are removed through random mortality.
The combination of pulsed resource supply and random removal of agents per round essentially qualifies this as a chemostat-type system. However, there may not be a simple connection between current continuous or discrete analytical formulations of chemostat systems and our simulator even under conditions of homogeneous mixing. This is because agents do not all reproduce in any given time step, or after consecutive time steps, which makes reproduction asynchronous among rounds, an added, realistic source of nonlinearity in system dynamics.
World description
The world specifies the combat matrix, which assigns pay-offs for agent-agent interactions, and it defines the transformation efficiency of resources by consumers. Resource exchange follows a series of steps in which a consumer agent determines whether to interact with a potential prey agent. The "decision-making" process is accomplished with the condition string. If a consumer agent's condition matches a prey agent's tag in whole or in part the consumer agent may choose to interact with the prey agent. Whether or not it will interact depends on the consumer's other choices of available prey. The actual interaction is controlled by the offense and defense strings. Once an interaction is instigated by one agent, each agent's offense string is pitted against the other's defense string to determine the amount of resources that are transferred from one agent to another. This can be a unilateral or bilateral exchange depending on the genetic coding and an element of chance.
The scoring functions describing the amount of resources exchanged between two interacting agents are encoded in the combat matrix (Table 1). The combat matrix is square with dimension (resource + 1). A resource letter in the defense string is read across the top, in the offense string down the side and a null character is the last term in the row and column. The actual score is calculated using the offense-defense string matches on a position by position basis. The individual scores for each offense-defense character are added to obtain a total. Each agent's relative score determines its percentage chance of "winning" that percentage of resources from the opponent. Either, both or neither may win.
For example, consider the genetic make-up of a hypothetical plant, herbivore and carnivore agent (Table 1). This is a simple case where chromosomes (tag, condition, offense, defense) are defined by only a few resource characters. Notice first that none of the different agent types have resource a in their interaction chromosomes. This is because only plants have the capacity to take-up a from the abiotic environment and convert it to resource b (plant carbon). In effect, all agents are composed of resource b and c.
In real world systems, individuals recognize one another as predator or prey or consepecific competitor, etc. Based on this recognition, individuals may choose to adopt certain behaviors leading to specific agent-agent interactions (e.g. prey may flee from predators, predators may subdue prey, competitors exploit a mutually beneficial resource). As a consequence of these behaviors, resources are exchanged unilaterally or bilaterally. All Echo-type simulators embody this recognition process to instigate agent-agent interactions.
For example, in Gecko a herbivore instigates an interaction with a plant agent to obtain resource b according to the following protocol. First, a herbivore agent's condition string is compared with a plant agent's tag string (Table 1). Both are b so the there is a match and interaction can proceed. The simulator next determines the percentage of resources to exchange (% pay-off) using values in the row and column of the combat matrix. First, take the value of the matrix corresponding to the resource character in the offense string of the herbivore and the defense string of the plant. to determine the herbivore's pay-off. Similarly, for the offense string of the plant versus the defense string of the herbivore to find the plant's pay-off. In the example (Table 1) the first comparison (offense herbivore against defense plant) gives a pay-off value of 0.15 indicating that 15% of the plant agent's resources can be lost to the herbivore. The second comparison (offense plant against defense herbivore) yields a pay-off of zero, thus the plant does not benefit from the interaction. Note also that this is not equivalent to specifying that 15% of the plant population is lost to herbivory each round. Once the amount to be exchanged is estimated, the likelihood of an exchange is estimated. The probability of successful exchange is estimated for each agent as
Each agent draws a random number against this probability. There is the potential here for plants to "escape" herbivory depending on the outcome of this random draw. Thus, we are able to specify the biologically realistic condition that a herbivore will not necessarily kill a plant agent. A plant agent only dies from herbivory if repeated herbivory or resource shortage causes the plant's resource pool to fall below the level required to pay the maintenance tax.
Carnivore-herbivore interactions result in the demise of prey as coded herein in the condition and tag strings of herbivore and carnivore agents (Table 1). Again, an interaction is instigated as the carnivore condition string matches the herbivore tag string. To determine the net amount of resource exchanged use the offense (defense) string of the carnivore and the corresponding defense (offense) string of the herbivore to obtain combat scores. The pay-off is calculated by selecting the matrix value corresponding to first character in the offense string (c) of the carnivore and defense string (c) of the herbivore and adding this to the matrix value corresponding to the second character in the carnivore's offense string (c) and the second character in the herbivore's defense string (Ø). The two matrix values are 0 + 1 = 1 indicating that carnivores will have a gross pay-off of 100% of the herbivore agent's resource pool. The opposite comparison (herbivore's offense string pitted against the carnivore's defense) yields a gross pay-off of zero for the herbivore. Again, the net resource exchange is unilateral. However, in this case the consumer takes all of the prey's resource pool and thereby kills the prey agent.
Individual prey agents will vary in the absolute amount of resources accrued in their storage pools. Thus individual consumers will vary in their absolute resource uptake depending on which individual prey they exploit. This nicely sets the stage for exploring the consequences of adaptive consumer behavior on trophic dynamics. It is possible to code a set of rules which specify that individual consumer agents should seek out and instigate interactions with the "fattest" prey agents within their neighborhood to maximize resource uptake rate and, correspondingly, survival and reproduction (fitness). This rule seems to contradict observations for many predator-prey systems in which predators usually only capture "weak and sick" individuals. However, in such cases the weak and sick usually yield the highest net pay-off given the costs associated with subduing individual prey items with different physical condition. In our simulator, the cost of subduing prey is identical for all prey agents so only the size of a prey agent's resource pool (its condition) determines the net pay-off.
Order of rule implementation during simulations
In each round, the rules for agent-agent interactions are applied in the following sequence.
1. Give everyone a chance to interact. First, the simulator constructs an instigator list. The number of times that an individual agent appears on this list represents its interactivity (i.e., whether it will attempt to consume one or more prey in a round). An instigator agent is then randomly selected from this list. Another agent (target) with whom the instigator will interact is selected from the ordered list of agents at a site. The choice of targets will depend on the instigator's neighborhood and the distance the instigator is able to travel. Each instigator has a fixed maximum movement distance, the search distance, and it is unidirectional. This is a simple strategy to avoid remaining in a location in which all prey have been exploited.
In this paper, we compare two different movement regimes for the instigator. The first, and simplest, is that the instigator's specified range is the mean of a negative exponential distribution. From this distribution, a distance is randomly selected and the instigator interacts with whatever it finds at that location. In the second, more sophisticated regime, the movement range is an inclusive search distance. Within this search distance, the instigator chooses a target with whom it "wants" to interact (perception). If there is more than one such target then the choice is predicated on specific rules for preference (see Simulating Trophic Dynamics below). If there is no suitable target within the search distance, the instigator moves the full search distance and interacts (or does not) with whatever agent is at the new location.
Once the instigator is in its new location then the condition string of each player is compared with the opponent's tag. If there is at least one match then an interaction takes place. Each agent pits its offense string against the opponent's defense string and the score is calculated using the combat matrix. All resources added to an agent via either combat or resource uptake from the site are added to the agent's "gullet" before resources are transformed. Because of inefficient resource transformation, it may take several meals before a consumer gathers enough of one resource type necessary to transform resources and allocate the new resource to the storage pool.
2. Distribute abiotic resources to autotrophic agents at the site. Resources are distributed to each agent that is capable of taking-up resources. They are allocated as equally as possible without exceeding an individual agent's uptake limit. The site retains excess resource up to the site maximum.
3. Take maintenance tax. A maintenance tax is collected from each agent. This tax rate is part of the agent's description. If an agent cannot pay its tax because its resource reserves are too low, the agent dies and its resources return to the site.
4. Kill agents at random. A fixed proportion of agents are randomly selected and killed each round. Victims of random mortality do not return their resources to the site. Typically, this cause of mortality is small relative to death rates due to interaction and taxation.
5. Add new resources to the site. New abiotic resources are added. The amount of resource added is specified by the site resource production function and cannot exceed the site maximum.
6. Migrate agents. Any consumer agent who did not receive a pay-off within the round tries to migrate to one of four or eight neighboring sites if there are any. This is a purely random assignment as the agent has no perception or choices; it is just "hungry" enough to test it's luck. New immigrants are ignored at their new location until the next round.
7. Self replicate. All agents are evaluated to determine if they have sufficient resources in their storage pools to reproduce. Any that can reproduce, do so. Newborns are inserted next to their progenitors. The newborn's complement of resources is deducted from the progenitor's resource pool. In addition, we transfer half the progenitor's remaining resource pool to the offspring (i.e., maternal effort). If sexual reproduction is enabled in the agent description, this takes place instead of self-replication, with the additional complication of finding a suitable mate.
Simulating trophic dynamics
We conducted a series of calibration experiments to evaluate qualitative dynamics of a 3-trophic level food chain comprised of a plant, herbivore and carnivore species. Because each simulation run of Gecko requires random selections, the simulator does not offer deterministic outcomes even when parameter values remain constant. The dynamics must, therefore, be interpreted in a probabilistic sense. For each run of the simulator, we defined stability as all species persisting for 1000 rounds. However, we required that the artificial food webs offer some semblance of reality by at least demonstrating known ecological properties, e.g., cascading trophic effects (sensu Paine 1980, Carpenter et al. 1986) and a trophic pyramid loosely defined as abundance of plants > herbivores > carnivores.
We began with a basic world in which there was (i) a 1:1 resource transformation efficiency (i.e., a = b = c) and (ii) random movement by heterotrophic consumers where the movement distance was drawn from a negative exponential distribution with a prespecified mean distance, e.g., 2, 4, etc. neighboring agents. This simulates a completely diffusive predator-prey system in which individual agents undergo homogeneous reciprocal or random "gas molecule" interactions when movement distance is unrestricted. The assumptions of 1:1 resource transformation and random encounters match those of conventional mathematical models of trophic dynamics in chemostat-type systems (Schoener 1973; Tilman 1982; Schmitz 1992). Under these assumptions, our simulator reproduced a persistent 3-level system. However, when we added realistic transformation inefficiencies, these dynamics broke down. We therefore explored which kinds of behaviors and realistic biological conditions required for stable dynamics. We found that persistence was facilitated by allowing individual heterotrophic agents the ability to make choices about resource selection (perception and intentional behavior).
Our implementation of perception and intention may be considered a rudimentary adaptive behavior. Individual agents are assigned the ability to perceive and choose the "fattest" prey agent, defined as the one with the largest resource pool, within the neighborhood. Exploiting the fattest prey agent in the environment is adaptive in the sense that this strategy yields the highest resource uptake per round. This increases the likelihood that an agent will survive after the resource tax is collected and allows an agent to reproduce sooner and more often.
Resource transformation is altered by specifying that there will be a 10% conversion efficiency of abiotic resources into plant biomass and a 33% conversion efficiency of plant resources into animal biomass (i.e. 30a = 3b = c). The exact choice of these parameter values is arbitrary but they are kept within the same magnitude as in realistic systems.
Basic world
In the basic world we allowed diffusive movement by agents. We obtained 4 alternative food web states depending on the degree and character of movement by agents in space (Fig. 2). Moreover, extensive simulations indicated that there were increasingly narrow ranges of parameter space in which higher levels of organizational complexity could be sustained. In essence, it was easier to have single or two populations to persist for a wide range of parameter values.
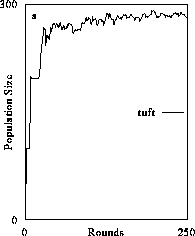
The first state (Fig. 2a) in which plants exist alone serves as a litmus test for internal consistency and qualitative congruence with existing analytic, mechanistic formulations of population growth. Abiotic resources are distributed as evenly as possible to all plant agents so the simulator matches the condition of egalitarian resource distribution in conventional mechanistic models (Schoener 1973, Schmitz 1992). In the absence of a consumer, plant populations reach an asymptote or carrying capacity. This is an emergent result of plants exploiting a pulsed supply of resources and not because there is a numerical maximum set. It can be determined quantitatively that carrying capacity is a function of resource supply rate, set by the resource production function, divided by resource demand for maintenance and replacement reproduction (maintenance tax). This emergent result is congruent with definitions of carrying capacity in analytic, mechanistic models of population and community dynamics which embody the assumption of chemostat-type consumer-resource dynamics (Schoener 1973, Tilman 1982, Schmitz 1992).
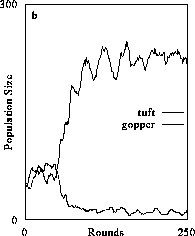
Higher levels of organizational complexity could be reached but only after movement strategies of consumers were altered. For example, the second state (Fig. 2b) was obtained when consumer movement was very localized such that resource exploitation is intense within a limited area. Carnivores were unable to exist in this system because herbivore populations could not reach sufficient densities to support them. Moreover, the carnivores were not sufficiently mobile to encounter prey at the rate needed to sustain their populations.
We obtained higher degrees of organizational complexity (Figs. 2c and 2d) once consumers became more mobile and essentially adopted a "mowing" strategy. "Mowing" here means eating (and moving) smaller meals more often, thus inflicting less damage over a greater range. With increased mobility we also saw an increase in the oscillatory behavior of consumer and resource populations. The effects of consumer mobility has also been explored by McCauley et al. (1993, called ambush vs. cruising strategies) in a diffusive system but on a lattice geography. They also observed a greater tendency for populations to oscillate when consumers adopt a cruising or mowing strategy. Consequently, the dynamics observed in our simulator are not an artifact of our particular topology.
The dynamical state in Fig. 2b was particularly curious. Essentially, plant populations exhibited damped oscillations to their carrying capacity and herbivores exist in a quasi steady state over 1000 time steps (the population may decline at a very low rate). At first glance, this state seemed highly counterintuitive because Lotka-Volterra models show that prey populations must fall below carrying capacity if they are being exploited by their consumers. However, this state makes perfect sense given the physiologically and behaviorally explicit properties of plant and herbivore agents and the spatially explicit nature of our simulator. Essentially, herbivores are only exploiting plants in localized spots and are consuming a proportion of the population equal in magnitude to the proportion that would normally have died from a failure to meet maintenance requirements. This idea was first introduced by Errington (1945) who called the phenomenon compensatory mortality. The idea has never received serious attention because the dynamic rarely if ever emerged in previous theoretical formulations of consumer-resource dynamics. Compensatory mortality arises in our simulator because plants are at carrying capacity, so their per capita resource reserves, by physiological definition, are only sufficient to meet maintenance and replacement reproduction. Thus, herbivores will only be able to exploit prey items with small resource pools which limits the rate at which they are able to transform plant resources into their own biomass. This, coupled with the fact that localized patches are being "spoiled" because the herbivores are not mobile, allows plants the capacity to balance the effects of herbivore exploitation at other locations where herbivores do not exist.
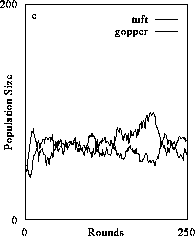
With increased mobility, herbivores were able to exert a strong limiting affect on plants and hold plants below carrying capacity (Fig. 2c). This state was, however, transitory as slight increases in movement enabled the system to shift rapidly to a full, three level food chain (Figs. 2d).
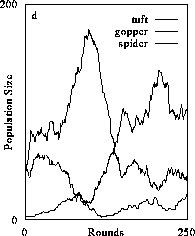
We observed persistence of the full, three level food chain (Fig. 2d) when consumers were completely diffusive. This result is congruent with analytical models of trophic dynamics which embody similar consumer-resource interactions (Schmitz 1992). However, we also had to adjust carnivore behavior to ensure persistence. Carnivores were extremely efficient at capturing prey agents and because of the 1:1 conversion of resources could translate resource intake into a high numerical response. This positive feedback in the carnivore population had a strong negative feedback on the herbivore population quickly leading to starvation of the carnivores. We decreased the positive feedback by making carnivores less efficient at prey capture. The easiest way to do this was to force consumers to become dormant for a specified number of rounds following ingestion of a meal. Basically, we decreased the probability that a consumer captured a prey item in a given time period. We recognize that this is one of many ways consumer behavior could be modified to stabilize dynamics. An exhaustive analysis detailing the consequences of a host of behavioral ecological strategies on trophic dynamics was beyond the scope of the current study. Nevertheless, our simulator offers considerable potential to explore such linkages.
Effects of consumer behavior
The assumption of diffuse movement was modified by allowing consumer agents the ability to perceive differences in resource quality among prey agents and direct their behavior to exploit the "best" prey agent within their neighborhood, a rudimentary adaptive behavior. The 3-level dynamics under these conditions is compared with case for random mixing in Fig. 3. Intentional consumer behavior causes the population levels of consumers and resources to converge. This is not because of increased numbers of consumer agents exploiting prey agents due to a higher consumer numerical response, but because there is increased capacity of consumers to limit their prey populations. Basically, consumers are eating those prey agents which are most likely to reproduce, thereby decreasing the reproductive capacity of the population. In these simulations, intentional behavior enhanced the persistence of the 3-level system because it was less likely that consumer-resource dynamics would decouple and drift to less complex organizational states (i.e., 2 or 1 species systems, Fig. 2b and 2a).
Effects of varying trophic efficiency
The effects of trophic efficiency in combination with perception and intentional behavior are presented in Fig. 4. Under simple diffusion (i.e., perception switched-off) the system never went beyond state 2 in parameter space (Fig. 4a). Essentially, this is because under the assumption of random gas-molecule mixing, consumers cannot find their prey in space through simple luck, due to the pyramid of numbers engendered by trophic inefficiencies. It was only after we introduced perception and intentional behavior that consumers were able to find their prey and so we were able to observe persistence of all three trophic levels (Fig. 4b).
Conclusions
Although the species composition and trophic linkages in the 3-level systems examined here can hardly be considered complex relative to the architecture possible in real-world systems, limiting our simulations to this level of complexity was deliberate. A full analysis of organizational complexity would have been premature without first introducing the simulator, demonstrating its potential and calibrating it to existing ecological systems.
Our simulator offers insight into some important mechanisms leading to sustained organizational complexity. In differential equation models, stability arises from negative feedback between consumer and resource populations or within populations. These interactions are expressed wholly at the population level. Our simulator shows that individual physiological capacity and behavior can have a profound effect on the character of higher-scale interactions. It is noteworthy that this is an emergent result of allowing individuals to interact according to simple biologically plausible rules (e.g., movement, prey choice, dormancy) rather than specify a priori the functional forms of interaction (e.g. functional response, higher order terms for density-dependence, etc.). We found that individual behavioral ecology of consumers could modulate the degree to which prey populations oscillated, decreasing the likelihood that a prey population would go to extinction. These results offer a different perspective on factors stabilizing food chain dynamics.
We have now set the stage to enlist the full potential of the simulator to explore systematically the effects of individual-level behavior on community-level dynamics with large numbers of interacting species--true food webs as opposed to simple food chains examined here. It is also possible to explore how natural selection on physiological and behavioral traits may shape the organizational structure of food webs by assigning agents differing resource transformation capabilities and different abilities to perceive resource quality.
Acknowledgments
We thank Günter Wagner, Leo Buss, John Holland, Stephanie Forrest, Ed McCauley, Andrew Beckerman and Kristina Rothley for helpful discussion and comments. Terry Jones kindly offered us his original implementation of Echo 0.1 upon which Gecko was built. The research was supported by NSF BIR-9400642, the Yale School of Forestry and Environmental Studies and the Yale Institute for Biospheric Studies. This is contribution # 24 of the Yale Center for Computational Ecology.
References
Abrams, P.A. (1990). Adaptive responses of generalist herbivores to competition: convergence or divergence. Evol. Ecol. 4:103-14.
Abrams, P.A. (1993). Indirect effects arising from optimal foraging. In Mutualism and Community Organization: Behavioral, Theoretical, and Food-web Approaches. (H. Kawanabe, J.E. Cohen, and K. Iwasaki eds.), pp. 225-79, Oxford University Press, Oxford U.K.
Brown, J.H. (1994). Complex ecological systems. In Complexity: Metaphors, Models, and Reality. (G.A. Cowan, D. Pines and D. Meltzer eds.), pp. 419-50, Addison-Wesley Publishing Co. New York.
Carpenter, S.R., J.F. Kitchell and J.R. Hodgson. (1985). Cascading trophic interactions and lake productivity. Bioscience 35:634-39.
Drake, J.A. (1990). The mechanics of community assembly and succession. J. Theor. Biol. 147:213-34.
Durrett, R. and S.A. Levin. (1994). The importance of being discrete (and spatial). Theor. Popul. Biol. 46: 363-94.
Errington, P.L. (1945). Some contributions of a fifteen-year local study of the northern bobwhite to a knowledge of population phenomena. Ecol. Monogr. 151:1-34.
Fryxell, J.M. and P. Lundberg. (1994). Diet choice and predator-prey dynamics. Evol. Ecol. 8:407-21.
Gell-Mann, M. (1994). The Quark and the Jaguar. W.H. Freeman, N.Y.
Hastings, A. (1990). Spatial heterogeneity and ecological models. Ecology 71: 426-28.
Hastings, A. (1993). Complex interactions between dispersal and dynamics: lessons from coupled logistic equations. Ecology 74:1362-72.
Holland, J. H. (1992). Adaptation in Natural and Artificial Systems. MIT Press, Cambridge.
Holland, J. H. (1994). Echoing emergence: objectives, rough definitions and formulations for ECHO-class models. In Complexity: Metaphors, Models, and Reality. (G.A. Cowan, D. Pines and D. Meltzer eds.), pp. 309-42, Addison-Wesley Publishing Co. New York.
Holt, R.D. (1983). Optimal foraging and the form of the predator isocline. Am. Nat. 122:521-41.
Ives, A.R. and A.P. Dobson. (1987). Antipredator behavior and the population dynamics of simple predator-prey systems. Am. Nat. 130:431-47.
Kim, J. (1992) Factors Influencing the Growth of Populations and Their Spatial Distribution. Dissertation, S.U.N.Y. Stony Brook.
Levin, S.A. (1992). The problem of pattern and scale in ecology. Ecology 73: 1943-67.
McCauley, E., W.G. Wilson, and A.M. DE Roos. (1993). Dynamics of age-structured and spatially structured predator-prey interactions: individual-based models and population-level formulations. Am. Nat. 142:412-42.
McLaughlin, J.F. and J. Roughgarden. (1991). Pattern and stability in predator-prey communities: how diffusion in spatially variable environments affects the Lotka-Volterra model. Theor. Popul. Biol. 40: 148-72.
Mitchell, M. and S. Forrest. (1994). Genetic algorithms and artificial life. Artificial Life 1:267-89.
Paine, R.T. (1980). Food webs: linkage, interaction strength and community infrastructure. J. Anim. Ecol. 49:667-85.
Post, W.M. and S.L. Pimm (1983). Community assembly and food web stability. Math. Biosci. 64:169-92.
Schmitz, O.J. (1992). Exploitation in model food webs with mechanistic consumer-resource dynamics. Theor. Pop. Biol. 41:161-83.
Schmitz, O.J. (1995). Functional responses of optimal consumers and the implication for regulation of resource populations. Wildlife Research 7(1): in press.
Schoener, T.W. (1973). Population growth regulated by intraspecific competition for energy or time: some simple representations. Theor. Pop. Biol. 4:56-84.
Schoener, T.W. (1993). On the relative importance of direct vs. indirect effects in ecological communities. In Mutualism and Community Organization: Behavioral, Theoretical, and Food-web Approaches. (H. Kawanabe, J.E. Cohen, and K. Iwasaki eds.), pp. 365-411, Oxford University Press, Oxford U.K.
Seymour, R.M. (1994). Optimal resource consumption, discrete dynamics, and intraspecific competition. Theor. Pop. Biol. 45: 132-66.
Tilman, D. (1982). Resource Competition and Community Structure. Princeton University Press, Princeton, N.J.
Wilson, W.G., A.M. DE Roos, and E. McCauley. (1993). Spatial instabilities within the diffusive Lotka-Volterra system: individual-based simulation results. Theor. Pop.Biol. 43: 91-127.
Wootton, J.T. and M.E. Power (1993). Productivity, consumers and the structure of a river food chain. Proc. Nat. Acad. Sci. (USA) 90:1384-87.
Yodzis, P. (1988). The indeterminacy of ecological experiments as perceived through perturbation experiments. Ecology 69:508-15.
Yodzis, P. (in press). Food webs and perturbation experiments: theory and practice. In Food webs : Integration of Patterns and dynamics.
| Combat Matrix | Genetic Makeup | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Defense | a | b | c | Ø | Plant | Herbivore | Carnivore | |||
| Offense | a | 0 | 0 | 0 | 0.1 | Tag | b | cc | c | |
| b | 0 | 0 | 0 | 0.1 | Condition | b | b | cc | ||
| c | 0.1 | 0.15 | 0 | 1.0 | Offense | b | c | cc | ||
| Ø | 0 | 0 | 0 | 0 | Defense | b | c | cc | ||
| Interactions | |
|---|---|
| plant vs. herbivore: b vs c = 0 | p(herbivore wins) = 0.15/(0.15 + 0) = 1.0 |
| herbivore vs. plant: c vs. b = 0.15 | |
| carnivore vs. herbivore: cc vs. c = 0 + 1.0 = 1.0 | p(carnivore wins) = 1.0/(1.0 + 0) = 1.0 |
| herbivore vs. carnivore: c vs. cc = 0 + 0 = 0 | |
Figures
(Figure 1 not available. Picture water fountain dripping letters on a necklace of pearls around its base. The pearls are white, gray, and black, randomly mixed.)
Figure 1. Diagrammatic representation of Echo and Gecko topology (modified after Holland 1992). Individual agents are arranged in a cyclic ordering with different agent types (e.g. plant, herbivore or carnivore) represented by different shaded circles. Abiotic resources are supplied as a pulsed input, a resource fountain of sorts, to autotrophic agents. (Return to discussion.)
|
|
|
|
Figure 2. The effect of diffusive consumer mobility on organizational structure of food chains composed of plants (tuft), herbivores (gopper) and carnivores (spider). Simulations producing results in (2a) consist only of plants which increased in abundance to reach a resource-determined carrying capacity. Introducing herbivores and varying their mobility (the number of agent positions over-which they could travel) results in increased organizational complexity because consumers were increasingly better able to exploit prey and gain sufficient resources to persist (2b and 2c). Above a certain level of exploitative ability, the herbivore population could sustain a third, carnivore trophic level (2d). (Return to discussion.)
 |
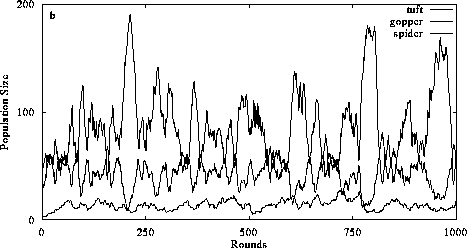 |
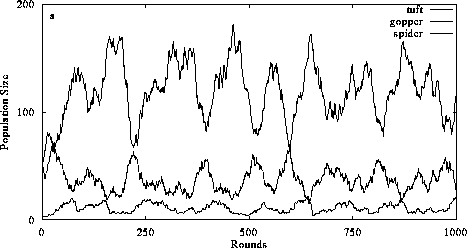
Figure 3. The effect of perception and intentional behavior on food chain dynamics. In 3a, perception is switched off and consumers exhibit diffusive mixing with prey. The addition of perception does not alter the organizational structure of the food chain (3b) but it increases the ability of consumers to limit their prey populations. (Return to discussion.)
 |
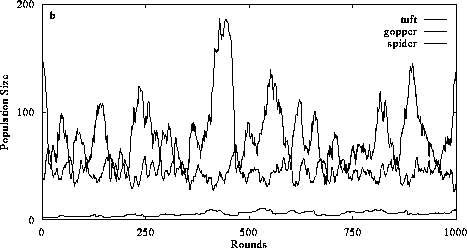 |
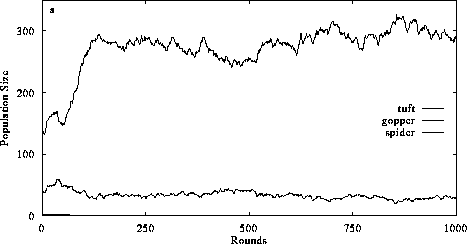
Figure 4. The effect of decreasing trophic efficiency and altering consumer perception on food chain structure. Introducing a 10% conversion efficiency of abiotic resources and 33% conversion efficiency of plant biomass into animal biomass in a system with diffusive mixing (4a) disrupts the organizational structure of the food chain (cf. Fig. 3a) primarily because herbivore equilibrium abundance decreases so carnivores are unable to gain sufficient resources to sustain their populations. Perception and intentional behavior (4b) increases the likelihood that the 3 trophic level system will persist, although carnivores occur in lower abundances than in systems with higher conversion efficiencies (cf. Fig. 3b). (Return to discussion.)